



Транскрипция, модель молекулярного уровня.
Классическую модель транскрипции рассмотрим по материалам, изложенным в статье “Лекция №5. Синтез ДНК, РНК и белков”. Лекция читалась в Московском физико-техническом институте на факультете молекулярной и биологической физики и этот материал должны все знать от студента до академика. Примерно, так интерпретируется режим транскрипции во всех учебных заведениях мира
Вот как описывается процесс транскрипции в данной лекции:
“Транскрипция – синтез РНК на ДНК, то есть синтез комплементарной нити РНК на молекуле ДНК осуществляется ферментом РНК-полимеразой. …у высших организмов (эукариотов) – несколько ферментов, они называются РНК-полимераза I, РНК-полимераза II, РНК-полимераза III… Каждый вид эукариотической РНК-полимеразы обладает своими специальными функциями, то есть транскрибирует определенный набор генов”.
Как же работает РНК-полимераза? Наиболее изучена РНК-полимераза кишечной палочки, ее работа и рассматривается.
“Бактериальная РНК-полимераза изучена очень подробно. Она состоит из нескольких белковых-субъединиц: двух α-субъединиц (это маленькие субъединицы), β- и β΄-субъединиц (большие субъединицы) и ω-субъединицы. Вместе они образуют так называемый минимальный фермент, или кор-фермент. К этому кор-ферменту может присоединяться σ-субъединица. σ-субъединица необходима для начала синтеза РНК, для инициации транскрипции. После того, как инициация осуществилась, σ-субъединица отсоединяется от комплекса, и дальнейшую работу (элонгацию цепи) ведет кор-фермент. При присоединении к ДНК σ-субъединица распознает участок, на котором должна начинаться транскрипция. Он называется промотор. Промотор - это последовательность нуклеотидов, указывающих на начало синтеза РНК. Без σ-субъединицы кор-фермент промотор распознать не может. σ-субъединица вместе с кор-ферментом называется полным ферментом, или холоферментом”.
И так мы видим, что для начала и продолжения транскрипции необходимо несколько белковых субъединиц: σ-, α-, α-, β-, β΄- и ω-. Причемσ-субъединицаработает только в начале процесса транскрипции, во время узнавания места, откуда начинается транскрипция. По сути, узнанное место это не что иное, как начало гена, который необходимо транскрибировать. Перед первым нуклеотидом транскрибируемого гена находятся либо нуклеотиды интронной части ДНК, либо стоп-кодон предыдущего гена. Как белковый комплекс различает эти нуклеотиды? Как он вообще оказывается всегда рядом с требуемым геном? А если он находится вдали от заданного гена, то по какому сигналу и каким механизмом доставляется в данное место? Холофермент - это конгломерат аминокислот. Конечно и человек - это конгломерат аминокислот и других молекул, и он может узнавать, двигать, удерживать и т.д. У человека все эти глаголы можно разложить на более мелкие действия. Свет падает на некий объект, объект отражает свет, фотоны света попадают в глаз человека, по зрительным волокнам попадают в мозг, создают там определенную энергетическую картину, котораясравнивается с ранее образованной картиной от такого же объекта. Если образы совпали, получаем соответствующий энергетический сдвиг поля мозга – узнали поле один сдвиг общего поля, не узнали – другой сдвиг общего поля. То же самое можно сделать и с остальными понятиями для человека. Но представить такое же явление для холофермента, это все равно, как представить, что наша галактика является таким же по свойствам холоферментом для вселенского великана. Это теория вложенности миров.
У человека около 30 000 генов и всего несколько типов полимераз. По каким признакам они делят между собой гены или они все работают одинаковым образом с каждым геном? Вопросов много, а ответов мало. Согласимся, что процесс узнавания произошел удачно. И что дальше?
“Связавшись с ДНК, а именно с промотором, который распознала σ-субъединица, холофермент расплетает двунитевую спираль и начинает синтез РНК”.
При репликации ДНК расплетается ДНК-хеликазой, а при трансляции холоферментом. Интересно чем отличается молекула, расплетенная ДНК-хеликазой, от молекулы, расплетенной холоферментом? Если различия есть, то в чем они заключаются? Данных о том, что в результате рас плетения к молекуле ДНК добавляется, или из нее изымается, какая-нибудь молекула, атом, электрон или ион обнаружить не удается. Поэтому объяснить различия в состоянии молекул с позиции молекулярной теории не представляется возможным. Эта разница прячется где-то на более глубоком уровне. Если различий нет, то почему не вступают в конфликт ДНК-полимераза и холофермент? ДНК-полимераза будет стремиться посадить на молекулу аденина молекулу тимина, а холофермент (иначе РНК-полимераза) будет старатьсясоединить аденин с урацилом. Кто кого пересилит и почему?
Раскручивание молекулы функция самой РНК-полимеразы, никаких топоизомераз в данном случае не требуется, для выполнения данной работы в комплексе имеется “шип”. Он и раскручивает молекулу. Описание устройства РНК-полимеразы предполагает, что с данной функцией она может справиться.
“Разберем более подробно молекулярную структуру кор-фермента. Как было сказано выше, кор-фермент состоит из α- и β-субъединиц. Они соединены так, что образуют как бы «пасть» или «клешню». α-субъединицы находятся в основании этой «клешни», и выполняют структурную функцию. С ДНК и РНК они, по-видимому, не взаимодействуют. ω-субъединица – небольшой белок, который также выполняет структурную функцию. Основная часть работы приходится на долю β- и β΄-субъединиц.
Внутри «пасти», которая называется главным каналом, находится активный центр фермента. Именно здесь происходит соединение нуклеотидов, образование новой связи при синтезе РНК. Главный канал в РНК-полимеразе – это то место, где во время элонгации находится ДНК. Еще в этой структуре сбоку есть так называемый вторичный канал, по которому подаются нуклеотиды для синтеза РНК.
Распределение зарядов на поверхности РНК-полимеразы обеспечивает ее функции. Распределение очень логично. Молекула нуклеиновой кислоты заряжена отрицательно. Поэтому полость главного канала, где должна удерживаться отрицательно заряженная ДНК, выложена положительными зарядами. Поверхность РНК-полимеразы выполнена отрицательно заряженными аминокислотами, чтобы ДНК к ней не прилипала”.
Несомненно, что «клешня» может развернуть молекулу, особенно в том случае, когда она знает, с каким шагом вести разворот. “Знает” должно содержаться в самом холоферменте, если он не получает сигналов из вне. Не понятно, где хранится это “знает”. Возможно в каком-то центре или в самой структуре. Структура холофермента для удержания молекулы приспособлена хорошо, лучше “клешни” ничего придумывать природе и не надо. Вместо зубцов на внутренней части “клешни” выложены положительные заряды и они притягивают к себе отрицательно заряженную молекулу, а в момент, когда надо продвинуть молекулу полярность внутренней части клешни изменяется, молекула отпускается и протягивается. Чтобы молекула все-таки держалась крепче, электростатические силы не особо надежны, ее прижимает “заслонка”, один из элементов РНК-полимеразы. Хотелось бы знать, какие силы и как продвигают молекулу в “клешне”. Об этом, к сожалению, ничего в лекции не сказано, но множество других параметров этого явления известны. Например:
“Элонгационный комплекс довольно стабилен, т.к. он должен выполнить большую работу. То есть, сам по себе он с ДНК не «свалится». Он способен перемещаться по ДНК со скоростью до 50 нуклеотидов в секунду. Этот процесс называется перемещение (или, транслокация)”.
Не раскрыта в лекции и работа вторичного канала, по которому подаются к активному центру нуклеотиды. Почему нуклеотиды, а не основания, рибоза и фосфат? Как все эти элементы оказываются в нужном количестве и в нужном порядке возле входа в канал? Если верно утверждение, что “Взаимодействие ДНК с РНК-полимеразой (кор-ферментом) не зависит от последовательности этой ДНК, в отличие от σ-субъединицы”, то может оказаться, что концентрация, например, цитозина возле входа в канал велика, а гуанина мала или гуанина вообще нет вблизи канала. Что же произойдет в данном случае? Машина транскрипции остановится, и будет ожидать подхода молекул гуанина? Но ведь это случайный процесс. Природа не может так поступать.
И что еще весьма странным кажется в модели транскрипции изложенной в лекции:
“Изменение конформации различных частей РНК-полимеразы происходит за счет изменения потенциальной энергии, что связано с электростатическими и гидрофобными взаимодействиями. Если молекулу РНК-синтазы «потрясти» (а «трясет» ее, также как и все другие молекулы в клетке, броуновское движение), то она начнет принимать конформацию с более низкой потенциальной энергией. То есть, источником движения молекулярной машины является энергия теплового движения отдельных ее составляющих, а устройство машины таково, что это движение приводит к нужному результату. При этом молекулярная машина потребляет энергию, которая, в основном, идет на изменение состояния тех или иных связей”.
Создается впечатление, что всю работу по транскрипции выполняет практически один комплекс в виде РНК-полимеразы, по крайней мере, для кишечной палочки. Никакие органоиды клетки в данном процессе не участвуют. Нет необходимости даже в источниках энергии в виде митохондрий. Достаточно установить соответствующий тепловой режим и молекулярная машина выполнит требуемую работу. Но это к счастью частный взгляд на процесс транскрипции. Эх, узнать бы что же это за устройство “таково”.
Из сайта “Молекулярная генетика: общие сведения” узнаем:
“Для процесса транскрипции генов требуются не только РНК-полимераза, но и другие белки, называемые основными факторами транскрипции. Установлено, что у эукариот таким фактором является ТАТА-связывающий белок (ТСБ, англ. ТАТА-Вох Binding Protein, TBP), который взаимодействует с основным регуляторным элементом. ТАТА-боксом, присутствующим в большинстве генов (см. с. 240). С этим комплексом затем связываются другие основные факторы транскрипции и РНК-полимеразы. Дополнительные факторы могут влиять на инициацию транскрипции, связываясь с другими регуляторными элементами. Отсюда они взаимодействуют с основным транскрипционным комплексом, либо активируя, либо ингибируя его”.
Как видим, для осуществления транскрипции требуются еще некоторые белки, т. е. продукты других, независимых генов. Как образуются эти белки, информация, о которых содержится в других независимых генах, никому не известно. Получается замкнутый круг: для синтеза белков требуются белки. И каждый из этих белков не приносится из вне, а синтезируется в каждой клетке. Эта коллизия не разрешима на уровне молекулярной генетики. В этой модели (несколько модифицированной модели “лекции…”) тоже не видно участия других органелл клетки в режиме транскрипции. Правда, приведены красочные схемы работы транскрипционного комплекса. Работа комплекса происходит примерно так.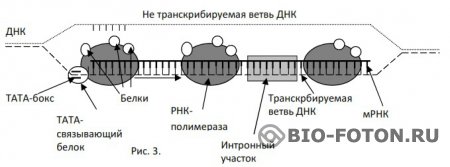
РНК-полимераза при помощи связывающего белка ТАТА (σ-субъединица) распознает код в ТАТА-боксе. Обычно это “Типичная последовательность (каноническая) — ...ТАТААА...”, но может быть и другой код, в этом случае РНК-полимеразе придется поработать больше, чтобы признать данный ген своим и начать синтезировать мРНК. В этом случае должны быть какие-нибудь второстепенные признаки для идентификации гена, чтобы не было ошибок. После распознавания комплекс присоединяется к ветви ДНК, начинает движение, и присоединяет к ней нуклеотиды, образуя информационную РНК. При встрече кода ...ААТААА... транскрипт отсоединяется от ветви ДНК. Дальше интронные части вырезаются, а экзонные куски сшиваются, и образуется линейная молекула белка. Кажется все логично, если бы не некоторые вопросы.
Что происходит на другой ветви ДНК во время транскрипции? Почему не вступает в работу ДНК-праймаза? Выше мы видели, что репликация может возникать в различных местах молекулы. Может быть эта ветвь ингибирована? Тогда что ее возбудит, чтобы после того как РНК диссоциирует от ДНК, ветви ДНК вновь соединились?
И самое важное. В каждой клетке организма существует полный набор генов. Их десятки тысяч в любом живом существе. Что в каждом гене содержится один и тот же (с небольшими вариациями) ТАТА-бокс? С наличием сайта полиаденилирования (обычно это последовательность ...ААТААА…) еще можно согласится, но представить один и тот же признак начала для всех генов не возможно. РНК-полимераза будет транскрибировать гены по случайному закону или в клетке будут параллельно происходить процессы трансляции, если в клетке несколько РНК-полимераз. А она должна точно знать, который из генов ей следует сейчас транскрибировать, ибо, если вместо коллагена синтезировать альбумин, получится нехорошо. Можно, конечно, их различать по ТАТА-боксу. Действительно последовательность ТАТАААТТАТАА… или другая представляют собой двоичный код (можно заменить Т на 1, а А на 0) и поэтому примерно 19 оснований в ТАТА-боксе могут присвоить личные номера каждому гену. Тогда надо предположить наличие внешнего указателя на требуемый код. Эту проблему нельзя объяснить в рамках молекулярной биологии. РНК-полимераза действует в пределах клетки и для нее все клетки равнозначны, и она не знает, для чего требуется данная клетка. Она как кирпич, который не знает, куда он ляжет - в стену жилого дома, забора, церкви или чего-нибудь другого.
Это только часть вопросов, на которые необходимо дать ответы молекулярной биологии. Естественно, что и в квантовой биологии будет не меньше вопросов, на которые не будет ответов, но это уже другие вопросы.